Les propriétaires de la US Corporation et les régisseurs de la fausse présidence de Biden semblent voués à l'échec après que leur plan de 150 000 milliards de dollars pour éviter la faillite ait été rejeté par les sociétés secrètes asiatiques. L'Occident fait maintenant face à une période de turbulences extrêmes alors que la mafia khazare se bat pour survivre avant que sa faillite ne devienne officielle début décembre, selon des sources du MI6 et de la société secrète asiatique.
Ce qui s'est passé, c'est que lorsque le KM a rencontré des représentants du gouvernement chinois dans un hôtel de l'aéroport de Zurich le 7 octobre (voir le rapport de la semaine dernière pour plus de détails), le KM a utilisé une combinaison de menaces et de promesses d'être « bon à partir de maintenant » pour acheter un encore quelques semaines. La partie chinoise a ensuite ramené à ses patrons une proposition de 5 000 milliards de dollars par an, sur 30 ans, de 150 000 milliards de dollars « pour lutter contre le réchauffement climatique ».
https://www.zerohedge.com/markets/here-hidden-150-trillion-agenda-behind-crusade-against-climate-change
Les Asiatiques ont été avertis par le MI6 que "le changement climatique est une opération de blanchiment d'argent, c'est l'effondrement de la dette souveraine américaine... c'est encore un autre prétexte pour cacher le fait que les familles de la Réserve fédérale sont en faillite". Bien sûr, les Asiatiques le savaient déjà et la proposition a été rejetée.
Cela peut être vu dans plusieurs reportages tels que les suivants du South China Morning Post :
Les dirigeants des nations les plus puissantes du monde se réuniront à Rome pour le Sommet du Groupe des 20, puis se rendront à Glasgow pour la Conférence des Nations Unies sur les changements climatiques (COP26) afin de cimenter les engagements climatiques avec d'autres pays.
Mais avec des informations selon lesquelles le président chinois Xi Jinping ne se rendra pas en Europe pour l'un ou l'autre événement, les observateurs occidentaux réduisent leurs attentes…
https://www.scmp.com/news/china/diplomacy/article/3151854/cop26-g20-prospects-dimmer-without-chinas-xi-jinping-room
Les Asiatiques sont intéressés par la préservation de la nature, mais sont également conscients qu'il n'y a aucune preuve scientifique que le carbone provoque le réchauffement climatique, selon une source de la société secrète asiatique. "Nous ne fonderons pas la politique mondiale de protection de l'environnement sur des mensonges", a expliqué la source.
"Nous devons résoudre les problèmes posés par la civilisation industrielle, maintenir les activités humaines dans les limites de l'écologie et de l'environnement, et mener une conservation holistique et une gouvernance systématique des montagnes, des rivières, des forêts, des terres agricoles, des lacs, des prairies et des déserts", a déclaré le président chinois. Xi Jinping explique.
http://www.news.cn/english/2021-10/16/c_1310247836.htm
Sans financement asiatique pour le cafouillage de la COP26, on s'attend à une grave crise en Occident -en particulier aux États-Unis- au cours des prochains mois. Cependant, les sources franc-maçonnes du MI6 et du P3 s'accordent à dire que -selon leur devise d'Ordo Ab Chao- (traduction "l'ordre du chaos") l'Occident doit sombrer davantage dans le chaos et le chaos avant qu'il ne soit enfin possible d'éliminer les derniers vestiges de Satanic Khazarian. règle mafieuse.
Déjà, près de 40 % des ménages américains ont été confrontés à de graves difficultés financières et n'ont pas pu se permettre de se nourrir et de se soigner, selon un récent sondage.
https://www.theguardian.com/us-news/2021/oct/12/us-households-financial-problems-medical-care-food
Ces commentaires publiés sur Zero Hedge montrent que ce n'est qu'un avant-goût :
Je distribue des vêtements très basiques, c'est-à-dire des t-shirts, des chaussettes, des sous-vêtements, etc. Magasins d'électronique à Portland Oregon… Toutes les entreprises avec lesquelles je fais affaire font une sorte d'augmentation rapide des prix.
https://www.zerohedge.com/personal-finance/are-you-prepared-mass-repricing-goods-and-services
Malgré les histoires sur le manque de chauffeurs de camion, etc., la vraie raison pour laquelle les Occidentaux, en particulier les Américains, manquent de choses, c'est parce que leurs chauffeurs d'esclaves KM sont en faillite.
Cependant, les sociétés secrètes asiatiques et occidentales ont déjà convenu en principe d'un plan pour ce qui se passera après la suppression du KM. Le plan présenté au Pentagone, au Commonwealth britannique, aux sages asiatiques et aux Russes est le suivant :
Planifier pour la version 2.0 des États-Unis
La United States Corporation est en faillite. Le Conseil de la Réserve fédérale de Saint-Louis et d'autres affirment qu'il a plus de 200 000 milliards de dollars de dettes et de passifs non financés. La dette a augmenté de façon exponentielle au cours des dernières années. Les tentatives pour lancer la boîte sur la route sont mathématiquement vouées à l'échec. Le seul choix est une déclaration formelle de faillite. Lorsqu'une entreprise fait faillite, même une grande entreprise, une nouvelle direction est amenée à prendre en charge les parties encore viables de l'entité en faillite.
Cependant, dans ce cas, il ne s'agit pas seulement de la faillite des États-Unis. Nous avons affaire à la faillite de familles aristocratiques majoritairement européennes qui possèdent l'ONU, la BRI, le FMI, la Banque mondiale, etc.
une alternative viable doit être mise en place pour prendre en charge le top management de toutes ces entités.
Considérons l'ONU. Les cinq membres permanents du Conseil de sécurité de l'ONU sont les États-Unis, la Russie, le Royaume-Uni, la France et la Chine. Une seule de ces nations n'est pas occidentale. Par conséquent, nous proposons de créer un nouveau conseil de sécurité composé de représentants de chacune des sept régions suivantes : Afrique, Amériques (Nord et Sud), Asie et Australasie hors Chine, Chine, Europe y compris Russie, Inde et monde islamique.
Les décisions seraient prises à la majorité et les veto ne s'appliqueraient qu'aux régions qui opposent leur veto. Un rôle spécial de droit de veto pour les Premières nations et les aînés est également envisagé.
Cet organe ne s'occuperait que des questions touchant la planète dans son ensemble. Les États-nations existants continueraient plus ou moins comme à l'heure actuelle.
Le conseil mondial de sept membres présiderait également une future organisation mondiale de planification (avec sept bureaux régionaux) qui prendrait en charge des projets d'amélioration du monde trop importants pour être réalisés par des États-nations individuels. Ceux-ci incluraient des choses telles que la fin de la pauvreté, l'arrêt de la destruction de l'environnement, la reconstitution des océans, l'augmentation de la biomasse mondiale totale et la colonisation de l'univers.
Les éléments fonctionnels et viables des organisations mondiales et régionales existantes seraient incorporés dans cette nouvelle structure.
En ce qui concerne les États-Unis, une nouvelle entité, basée à l'extérieur de Washington DC (peut-être l'Idaho, le centre géographique de l'Amérique du Nord) assumerait un rôle de gardien pour maintenir les éléments vitaux du régime américain actuel (police, trafic aérien contrôle, l'armée, les hôpitaux, etc.) en cours d'exécution.
Dans ce scénario, les militaires américains et canadiens utiliseraient le système de diffusion d'urgence et les réseaux médiatiques existants pour informer la population de la faillite de la société américaine. Ils expliqueraient que la faillite et le fait que les gouvernements civils au Canada et aux États-Unis avaient été pris en charge par des gangsters étaient les raisons de la déclaration d'urgence. Un régime intérimaire serait dirigé par l'armée et le personnel sélectionné du gouvernement et des entreprises existantes pour présider au rétablissement d'une démocratie viable, d'une presse libre et de la méritocratie. Tout se ferait de manière totalement transparente avec un retour constant des gens.
Le régime provisoire ne serait pas responsable des dettes de la société en faillite.
À une date ultérieure, une union avec l'Amérique latine serait également négociée.
Quant à l'Europe, les 47 nations du Conseil de l'Europe serviraient de base à une nouvelle sorte d'union régionale à négocier par les nations concernées. La Russie et l'OTAN fourniraient des garanties militaires.
Les autres régions mentionnées ci-dessus prendraient leurs propres décisions sur la façon de procéder.
Cette proposition a été acceptée comme base de négociations détaillées qui seront élaborées par les dirigeants mondiaux au cours des prochains mois, selon des sources du MI6, de la franc-maçonnerie P3 et des sociétés secrètes asiatiques.
Pendant ce temps, le monde est confronté à un danger et à un chaos extrêmes alors que le KM se bat bec et ongles pour sa survie. Les signes du chaos, bien sûr, se multiplient de façon exponentielle dans la plupart des pays occidentaux alors que le KM tente de rester au pouvoir en imposant sa marque au-delà du totalitaire de la bête Certificate Of Vaccination IDs (COVID).
Inutile de dire que des fuites de dénonciateurs de KM, des grèves de travailleurs, des arrestations massives, etc. font que ce plan visant à imposer un esclavage perpétuel et universel à l'humanité est voué à l'échec.
Le reportage lié ci-dessous est un exemple :
Les boîtes de vaccin COVID-19 d'AstraZeneca indiquent une date de fabrication du 15 juillet 2018. . . Mais "COVID-19" n'a été découvert qu'en 2019 et n'a été nommé que le 11 février 2020.
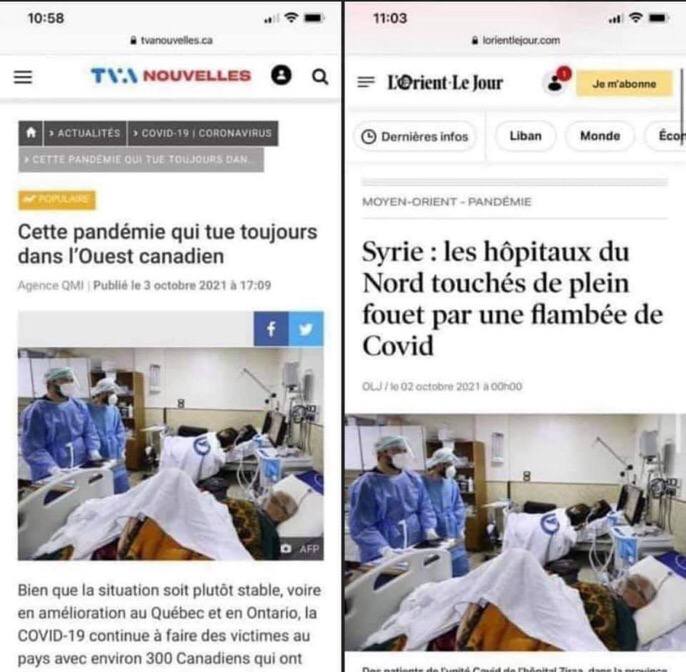
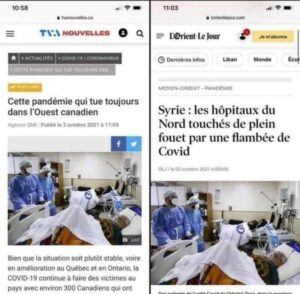 Les hôpitaux canadiens et syriens sont parfaitement identiques ! Les mêmes soignants, le même matériel, et même les patients sont identiques
Les hôpitaux canadiens et syriens sont parfaitement identiques ! Les mêmes soignants, le même matériel, et même les patients sont identiques
https://halturnerradioshow.com/index.php/en/news-page/world/how-did-astra-zeneca-manufacture-covid-19-vaccine-in-july-of-2018-before-the-disease- a-même-découvert-ou-nommé
Les images suivantes montrent un peu plus des informations qui sortent.
Tout cela signifie qu'il y a eu un "changement radical au sein du complexe militaro-industriel", selon des sources du MI6. « Toutes les polices du monde et toutes les agences de renseignement du monde en ont marre, nous en avons assez. Nous arrivons au point où nous disons « faites-les tous sauter ». La grande majorité du monde des célébrités sont des violeurs. Nous avons des preuves sur chacun d'eux et ils le savent. La plupart d'entre eux sont morts, ce que nous voyons sont des doubles du corps. Lorsque nous publierons nos informations, elles auront toutes disparu. Nous continuerons à nous battre très fort. »

Ce combat redevient également personnel pour votre correspondant. Munakata Hisao, un médecin de 73 ans qui prononce des discours pour les mêmes personnes qui organisent des événements pour moi, est décédé dans des circonstances mystérieuses. Dans ses conférences, il a expliqué en détail pourquoi les vaccins étaient dangereux et la pandémie suspecte. Fin août, il est tombé malade après un engagement public. On lui a dit de passer un test PCR mais il a refusé. Plus tard, il a été retrouvé inconscient à son domicile et emmené à l'hôpital. Là, il refusait toujours de passer un test PCR et utilisait une herbe chinoise appelée Ara. Puis il s'est amélioré. Alors qu'il était sur le point de quitter l'hôpital, son état s'est soudainement aggravé et il est décédé le 29 septembre. En outre, six personnes qui ont assisté à sa conférence d'août sont également tombées malades et sont décédées. Même si le médecin a refusé de passer un test PCR, sa mort a été répertoriée comme due à COVID 19.
Un autre de mes collègues qui a donné une conférence avec le Dr Munakata dit qu'il a eu une diarrhée sévère en même temps. Il prétend s'être guéri en jeûnant. C'est exactement ce que j'ai fait lorsque j'ai eu la diarrhée après avoir mangé une omelette sur mon vol d'Air Canada à mon retour du Canada à la fin du mois d'août. Plus tard, quelqu'un a sonné à ma porte en prétendant rechercher les symptômes du COVID et a semblé surpris de me trouver en vie.
Le 14 octobre, j'ai été averti que des gangsters dans une Rolls Royce avec la plaque d'immatriculation Shinagawa 311x Tsu 11 (品川311xつ11) m'attendaient en embuscade à l'extérieur d'un événement de prise de parole en public que j'ai organisé.
Le 10 mars 2011, la fille de Masaru Takumi
https://en.wikipedia.org/wiki/Masaru_Takumi
le surveillant financier du plus grand gang du Japon, le Yamaguchi Gumi (qui a été tué en 1997) a insisté avec une grande urgence pour que je la voie. Quand je l'ai rencontrée, elle m'a dit qu'on lui avait demandé de régler l'alarme sur mon téléphone pour qu'il sonne à 11 heures du matin le 11 mars, le jour du tsunami. On m'a également enveloppé d'un tissu autour de la taille « contenant des plantes médicinales » et on m'a dit de le garder pendant 24 heures. J'étais presque sûr qu'il contenait du poison et je l'ai enlevé dès que je suis rentré à la maison. Alors qu'en pensez-vous, est-ce une coïncidence qu'une voiture portant le numéro 311x tsu 11 m'attende ?
Lorsque j'ai dirigé cet incident par divers gangsters et membres de droite proches de l'empereur, ils ont dit que ces personnes sous-traitaient pour les Rockefeller et les Rothschild. "Ils sont en colère parce que [l'ancien ministre des Finances Heizo] Takenaka a perdu la capacité de piller le fonds de pension japonais pour eux", a expliqué un homme de droite. Une contre-attaque est maintenant en cours. La plupart de la pègre japonaise ne fait pas partie de la masse salariale de KM et se bat contre les gangsters qui sous-traitent toujours pour eux.
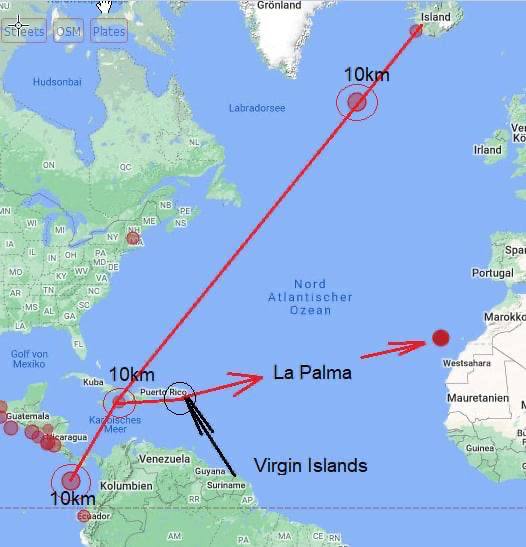
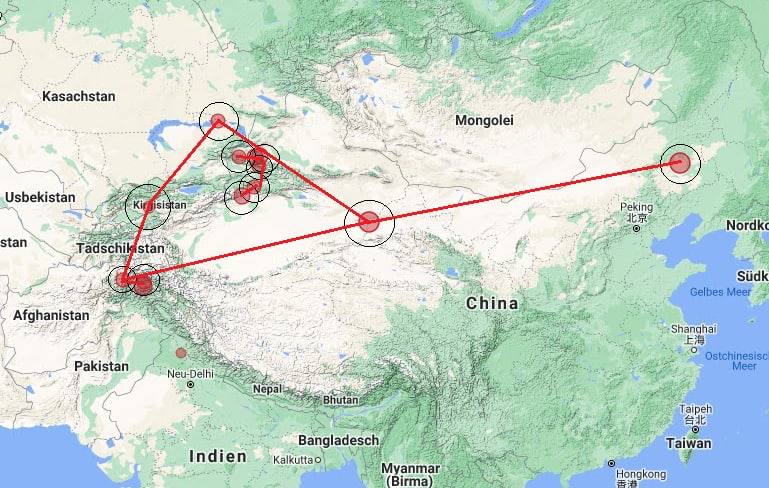
Sur une note finale cette semaine, nous recevons de nombreux rapports sur la force spatiale secrète détruisant des bases souterraines autour de la planète en vue de la "libération planétaire". Par principe, je m'en tiens au monde visible ici/maintenant. À ce niveau, nous pouvons certainement voir qu'il y a eu beaucoup de tremblements de terre dans le monde qui se sont produits au même épicentre à 10 kilomètres sous le niveau du sol et qu'ils se déroulent en ligne droite.
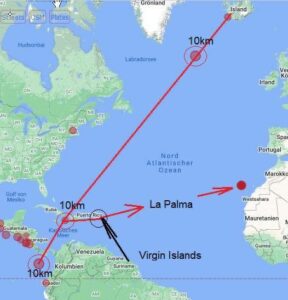
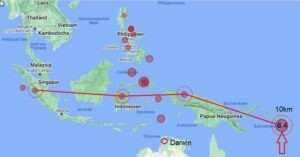
Il est peut-être vrai que les réseaux de tunnels souterrains sont détruits. Nous verrons. En attendant, la bataille pour la planète Terre se poursuivra jusqu'à ce que l'humanité soit enfin libérée de millénaires d'esclavage.